Livets opprinnelse -Oparins hypotese
(Fra boka 'The Design of Life' kap 8.4; av W. Dembski og J. Wells)
I 1924 foreslo den russiske biokjemiker Alexander Oparin en rent materialistisk tilnærming for livets opprinnelse, som har satt kursen for opprinnelse-til-livet forskning siden da. (1) I følge ham oppsto den første cellen av ikke-levende materie, ikke alt på en gang, men gradvis i stadier. Oparin holdt for at enkle kjemikalier slo seg sammen og dannet organiske sammensetninger som aminosyrer. Disse kombinerte seg så i neste omgang for å danne store, komplekse molekyler, som proteiner. Og disse hopet seg opp og formet sammenhengende nettverk innenfor cellevegger.
Bilde 1. Jordas nåværende atmosfære
 I følge Oparin var atmosfæren på den tidlige jorda veldig forskjellig fra den nåværende. Energikilder som hete fra vulkaner eller lynnedslag, ble sagt å virke på rene karbon-sammensetninger i atmosfæren, og forvandle dem til mer kompliserte sammensetninger. I den tidlige jordas sjøer, ble disse nylig dannede sammensetninger sagt å forme mikroskopiske masse-klustre, forløperne til tidligste levende celler på jorda.
I følge Oparin var atmosfæren på den tidlige jorda veldig forskjellig fra den nåværende. Energikilder som hete fra vulkaner eller lynnedslag, ble sagt å virke på rene karbon-sammensetninger i atmosfæren, og forvandle dem til mer kompliserte sammensetninger. I den tidlige jordas sjøer, ble disse nylig dannede sammensetninger sagt å forme mikroskopiske masse-klustre, forløperne til tidligste levende celler på jorda.
Oparin la således grunnlaget for en teori for prebiotisk eller kjemisk-evolusjon, som hevdet livet startet i en sjø av kjemikalier, ofte kalt prebiotisk suppe. Til tross for mange modifikasjoner, har denne hypotesen dannet den evolusjonære standardmodellen for livets opprinnelse. I følge Oparins hypotese vil ikke endring alene kunne styre nødvendige kjemikalier og sammensetninger som trengs for å danne komplekse biomolekyler og derfra til livet. I stedet hevder hypotesen en internt iboende tendens ved materien mot selvorganisering, som gir opphav til de ordnede strukturene vi ser i livet. Vi skal se på denne hypotesen mer detaljert, og spesielt drøfte forutsetningene den bygger på.
Forutsetning 1. Reduserende atmosfære. Jordas tidlige atmosfære inneholdt rikelig med fri hydrogen, og lite eller ikke noe oksygen.
Kommentar:
Alle eksperiment som simulerte atmosfæren til den tidlige jorda, har ekskludert fri oksygen. Det er fordi oksygen opptrer som et verktøy i ei girkasse, og aktivt hindrer kjemiske reaksjoner som produserer organiske komponenter. Dersom noen slike komponenter skulle ha blitt dannet, ville fri oksygen raskt ødelegge dem i oksiderings-prosessen. Som følge av dette har standard-teorien for kjemisk evolusjon antatt at det ikke fantes oksygen i jordas atmosfære, ved livets opprinnelse. Likevel har geologer nå sterke bevis for at betydelige mengder av oksygen var til stede i jordas atmosfære fra tidligste tider -da livet ble til. (2) F.eks. reagerer mange mineraler med oksygen -sml. jernoksid (rust), og resulterende oksider er funnet i stener som er eldre enn livets opprinnelse.

Om oksygen var til stede i den tidlige jordas atmosfære, så ville det ha vært umulig for organiske komponenter å ha blitt dannet og akkumulert, slik det skjedde i Urey-Millers eksperiment (1953). Likevel er det et tankekors at uten oksygen, kunne kanskje heller ikke organiske komponenter ha blitt bevart. Det ville vært nødvendig med betydelige mengder av oksygen, for å danne det beskyttende ozon-laget som beskytter jorda mot livsfarlig ultra-violett stråling. Siden livet blomstret på den tidlige jorda, så kan en realistisk simulering av jordas tidlige atmosfære trenge å inneholde oksygen.
Bilde 2. Oppsett til Urey-Miller forsøket
Et annet problem er at Oparins hypotese, og Miller-Urey eksperimentet, forutsatte at den tidlige jords atmosfære var hydrogen-rik. Likevel konkluderte geo-kjemikere på 1960-tallet at jordas opprinnelige atmosfære, kun inneholdt sporbare rester av hydrogen (H). Fordi jordas tyngdekraft var for svak til å holde på H, ville mesteparten av den forsvinne ut i rommet. Uten H til å reagere med CO2 og Nitrogen (N), kunne ikke metan og ammoniakk ha vært vesentlige komponenter i den tidlige atmosfæren.
Hva om Miller-Urey eksperimentet ble gjentatt med en mer realistisk blanding av vanndamp, CO2 og N? I 1983 rapporterte Miller at han og kolleger var i stand til å produsere en liten mengde av den enkleste aminosyren glysin, men det var det beste de kunne få til i fravær av metangass. I 1991 summerte John Horgan opp status for forskningen: en atmosfære av CO2, N og vanndamp, ville ikke bidratt til dannelse av aminosyrer. (3)
Forutsetning 2. Enkle organismer som dannet ursuppa, ble på et vis bevart, slik at energien ansvarlig for å danne dem ikke også ødela dem.
Kommentar: Noen kjemiske reaksjoner skjer spontant, i andre motstår stoffene hverandre. Å drive fram en slik reaksjon, krever energi, i form av varme, f.eks. elektrisitet. men og her er et viktig paradoks, energi bryter også kjemiske komponenter i stykker. Det er dermed vitalt at de destruktive effektene av energi, ikke overgår de konstruktive ved livets opprinnelse. Men når vi tar de destruktive effektene av energi på den tidlige jorda i betraktning, så ville det favorisere enkle framfor komplekse molekyler, som ikke har noen egen kapasitet til spontant å organisere seg selv til maskineriet i en levende celle.
 Bevisene for at den tidlige jorda bevarte enkle organiske materialer, er knapt overbevisende. Atmosfæriske simulerings-eksperimenter neglisjerer destruktive effekter av energi. I slike eksperimenter er aminosyrer og andre organisk relevante produkter skilt ut ved en 'felle', der de beskyttes fra de destruktive kreftene ved elektriske ladninger. Men om de hadde blitt utsatt for slike utladninger på den tidlige jorda, ville de ha løst seg opp så snart de ble dannet. Slike feller som benyttes i forsøk, korresponderer ikke realistisk med noen beskyttende mekanisme, som fornuftig antas å ha eksistert på den tidlige jorda.
Bevisene for at den tidlige jorda bevarte enkle organiske materialer, er knapt overbevisende. Atmosfæriske simulerings-eksperimenter neglisjerer destruktive effekter av energi. I slike eksperimenter er aminosyrer og andre organisk relevante produkter skilt ut ved en 'felle', der de beskyttes fra de destruktive kreftene ved elektriske ladninger. Men om de hadde blitt utsatt for slike utladninger på den tidlige jorda, ville de ha løst seg opp så snart de ble dannet. Slike feller som benyttes i forsøk, korresponderer ikke realistisk med noen beskyttende mekanisme, som fornuftig antas å ha eksistert på den tidlige jorda.
Bilde 3. Moderne analogi til 'ursuppa'
Forutsetning 3. Konsentrasjon. Biologiske sammensetninger akkumulerte til tilstrekkelig høye konsentrasjoner, slik at de kunne kombinere med hverandre for å danne de komplekse molekylene, som er nødvendig for livet.
Kommentar:
Mange reaksjoner som er nødvendige for å danne viktige biologiske sammensetninger er blitt observert under, og ofte bare under, kunstige laboratorie-forhold. Samtidig virker mange naturlige reaksjoner mot dannelse av biologisk viktige sammensetninger. F.eks. reagerer ikke aminosyrer spontant med hverandre. De reagerer imidlertid raskt med andre substanser, som sukker. Men det innebærer at om aminosyrer ble dannet på den tidlige jorda, så ville de ikke bare ligge og flyte i sjøer og vente på at de rette aminosyrene dukket opp. I stedet ville de reagere med andre sammensetninger i alle slags kryss-reaksjoner, bundet og utilgjengelige for noen biologisk nyttig funksjon. I prebiotiske eksperimenter er dominerende resultater store mengder av ikke-biologisk 'mudder'.
For at Oparins hypotese for livets opprinnelse skulle holde, ville enkle biologiske komponenter måtte konsentreres i tilstrekkelige mengder, før de kunne kombineres til å danne store komplekse molekyler som er nødvendige for livets opprinnelse. Naturen opererer ikke med noen spare-plan for å sette til side biologisk viktige sammensetninger for evt. fremtidig bruk. Ingen bio-polymerer er dannet under eksperimenter som simulerer primitiv atmosfære, unntatt noen meget små peptider. Det meste av det som er produsert i slike eksperimenter, er som nevnt hydrogen-fattige uoppløselige materialer, ikke-biologisk slam. (4) Selv under de mest favoriserende forhold, skjer forstyrrende kryss-reaksjoner og kaster tvil over Oparins hypotese om 'konsentrasjon'.
Forutsetning 4. Uniform orientering. Bare L-amino-syrer kombinerte seg for å produsere livets proteiner, og bare D-sukker reagerte for å danne poly-sakkarider og nukleotider.
 Kommentar:
Aminosyrer, sukker, proteiner og DNA er ikke bare kjemikalie-sammensetninger. De fremviser meget spesifikke tredimensjonale strukturer. Når de syntetiseres i laboratoriet, kan de ha de rette bestanddeler og likevel inneha feilaktig tre-dimensjonal form. F.eks. opptrer aminosyrer i to ulike former eller kiraliteter. Disse er speilbilder av hverandre, og de to formene refereres som (høyre) D-form og (venstre) L-form. I eksperimenter og i naturen, forekommer de i 50-50 blanding. Levende organismer innehar bare L-formen av aminosyrer, uten at noen vet hvorfor. Om bare én D-form av aminosyre slapp til i et protein, så ville proteinets evne til å fungere minke, og ofte bli fullstendig ødelagt.
Kommentar:
Aminosyrer, sukker, proteiner og DNA er ikke bare kjemikalie-sammensetninger. De fremviser meget spesifikke tredimensjonale strukturer. Når de syntetiseres i laboratoriet, kan de ha de rette bestanddeler og likevel inneha feilaktig tre-dimensjonal form. F.eks. opptrer aminosyrer i to ulike former eller kiraliteter. Disse er speilbilder av hverandre, og de to formene refereres som (høyre) D-form og (venstre) L-form. I eksperimenter og i naturen, forekommer de i 50-50 blanding. Levende organismer innehar bare L-formen av aminosyrer, uten at noen vet hvorfor. Om bare én D-form av aminosyre slapp til i et protein, så ville proteinets evne til å fungere minke, og ofte bli fullstendig ødelagt.
Forskere har forsøkt via eksperimentering å bestemme hvordan preferansene kan ha oppstått ved materielle mekanismer i levende organismer. Resultatet er i beste fall en liten disproporsjonalitet av de to formene i forhold til hverandre. På dette og andre områder, viser livet karakteristika som er fremmed for noe som er kjent å produseres under naturlige forhold.
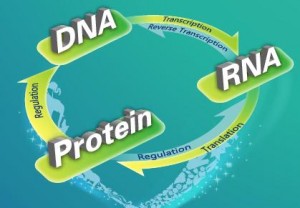
Bilde 4. Gjensidig avhengige: DNA-RNA-proteiner
Forutsetning 5. Simultan oppdukking av interlineære bio-makromolekyler (DNA og proteiner). Det genetiske maskineriet som forteller cellen hvordan danne proteiner og nødvendige proteiner for å danne det genetiske maskineriet oppsto gradvis og var nærværende og fungerende i de første fungerende protocellene.
Kommentar:
Det er når aminosyrer av rett kiralitet er konsentrert på rett plass, at det virkelige arbeid med å konstruere livet starter: Aminosyrer av L-formen trenger å kombineres i rett sekvens for å danne proteiner og nukleotidbaser knyttet til et (D-)sukker-fosfat rammeverk trenger å kombineres i rett sekvens for å danne DNA og RNA. Et vanlig protein er flere hundre aminosyrer langt, og DNA kan millioner av nukleotid-baser, som fortrinnsvis må være i rett sekvens.
Å syntetisere polymerer innebærer to ting: a) få tilhørende monomerer å slutte seg skikkelig sammen og b) få disse til å danne en funksjonelt meningsfull orden. Aminosyrer kan slutte seg sammen på ulike vis, og danne varierende kjemiske bånd. Likevel er det bare ett nødvendig kjemisk bånd, som kalles peptid-bånd, som inntreffer i funksjonelle proteiner. Arrangementet av aminosyrer i proteiner, er lineært -og vil ikke fungere om det er forgrenet. For å sammenlikne med en sekvens av bokstaver, er det mulig at bokstaver kan stå opp-ned, i ulike skråstillinger etc, men er bare entydig lesbare om de står rett opp og ned. På samme måte med dannelse av DNA og RNA-lenker, så må de kobles som '3-5' koblinger, selv om '2-5' koblinger dominerer utenom cellulært liv. Uten katalysator for peptid-bånd i aminosyre-sekvenser, eller '3-5' fosfor-lenker for nukleotid-sekvenser, kan det ikke være noen materialistisk vei til proteiner, DNA og RNA. Men de eneste katalysatorene vi kjenner, som kan håndtere denne oppgaven er enzymer og andre protein-baserte produkter, og disse forutsetter igjen hele DNA-RNA-protein maskineriet. Men det er nettopp dette maskineriet opprinnelse-til-liv forskningen skal forklare.
Vi har altså sett at de kjemiske kjente veiene konspirerer mot en naturlig dannelse av funksjonelle polymerer, som proteiner og DNA.
 Forutsetning 6. Funksjonell integrasjon. De høyst organiserte arrangement av tusener av deler i cellens kjemiske maskineri, som trengs for å oppnå cellens spesialiserte funksjoner, oppsto gradvis i kolloid-rik viskositet eller andre protoceller.
Forutsetning 6. Funksjonell integrasjon. De høyst organiserte arrangement av tusener av deler i cellens kjemiske maskineri, som trengs for å oppnå cellens spesialiserte funksjoner, oppsto gradvis i kolloid-rik viskositet eller andre protoceller.
Kommentar:
Om en har et reagensrør med steril saltoppløsning, og plasserer en levende celle inni dette -som en så stikker hull på med en steril nål. Da har man tilgjengelig alle molekyler i en levende celle, i rette forhold under rette betingelser, og uten kryssvirkende reaksjoner. Men uansett anstrengelser, vil ikke molekylene danne overgangen fra ikke-liv til liv. Og om teknologien noensinne skulle kunne sette sammen 'lille-trille' igjen, ville det ikke være bevis på selv-dannelse, men på design.
Bare for å bygge ett enkelt protein i en celle, trengs ca. 60 proteiner som fungerer som enzymer. De 60 proteinene trenger å samhandle med cellens genetiske DNA-maskineri. Om et eneste av disse enzymene mangler, vil ikke cellen være i stand til å danne proteiner. Enzymene involvert i proteinsyntese, er dermed uunnværlige for livet og samlet betraktet, danner de ett ikke-reduserbart komplekst system. De 60 enzymene må koordineres til å være samlet til stede på samme ørlille område i cella. Men slik koordinering forutsetter et enormt komplekst kommunikasjons-, transport og kontroll-system, som i seg selv er protein-drevet. Dermed faller Oparins forutsetning at denne oppbygning kunne skje gradvis ved materialistiske prosesser.
Bilde 5. Komplekse cellemembraner i nåværende celler
Forutsetning 7. Fotosyntese. De kjemiske prosessene, kjent som fotosyntese som opptar, lagrer og benytter solenergien til å danne næringsstoffer, oppsto gradvis innen kolloid-rik viskositet.
Kommentar: Intet nåværende kjente materialistiske mekanismer gjør rede for hvordan en så kompleks prosess som fotosyntesen (se Origo nr. 142) kunne ha blitt dannet. Biologer har spekulert mye i hvordan det kan ha skjedd, men disse spekulasjonene er ikke testbare. (5) De fortreffelige fininnstilte fotosyntetiske kompleksene for å fange sollys effektivt ved å utnytte kvante-teoretiske effekter og så ved å overføre den energien til reaksjonssentre i cellen, der de kan lagres og senere benyttes, vitner om et design-nivå som overstiger noe som er produsert så langt innen feltet av fotokjemisk ingeniørvitenskap. (6)

Gitt den sannsynlige sammensetning av den primitive atmosfære, spesielt med fri oksygen og ikke fri hydrogen, favorisering av enkle kontra komplekse molekyler samt nærværet til kryss-agerende interaksjon og fravær av homo-kiralitet utenom livet, og funksjonell integrasjon til f.eks. fotosyntesen, innenfor cellemembraner -så er det ingen grunn til å mene at forholdene på den tidlige jorda favoriserte oppdukking av livets byggeklosser ved rent naturalistiske krefter. Det mest sannsynlige tidlig-jord scenario, er ikke ett som bygger biologisk kompleksitet, men ett som bryter det ned. Det er verken en solid teori eller lovende eksperimentell basis for at bio-makromolekyler kunne ha vært dannet ved naturkrefter alene, mye mindre at en samvirkende og koordinert samling av dem, gradvis kunne ha utviklet seg til fungerende enheter med cellers kompleksitet.
Bilde 6. Oppsummering om livets opprinnelse
Dermed synes samtlige av Oparins forutsetninger angående den tidlige jords atmosfære og evne til å danne liv, ikke å være korrekte.
Referanser:
1.A. I. Oparin, The Origin of Life on Earth, 3.utg. New York: Academic Press, 1957
2. J.H. Carver, "Prebiotic Atmospheric Oxygen Levels," Nature 292 (1981): 136-138; J.F. Kasting, "Earths Early Atmosphere," Science 259 (1993): 920-926
3. John Horgan, In the Beginning, Scietific American 1991: 116-126
4. R. Shaphiro, "A Simpler Origin of Life", Scientific American (Febr.12, 2007):
5. "Photosyntesis Analysis Shows Works of Ancient Genetic Engineering." Science Daily (Nov.22, 2002)
6. Gr. S. Engel et al, "Evidence for Wavelike Energy Transfer through Wuantum Coherence in Photosyntetic Systems," Nature 446 (12. april 2007): 782-786
Oversettelse og bilder ved Asbjørn E. Lund
{kind=link}